|

Nobel Prize Design in DNA IV
Function, Specificity, and Complexity:
 The discovery of split genes brought Richard Robert and Phil Sharp the [1993]Prize. This came late in the examination of DNA because most early DNA research was done on bacteria which do not have split genes. In multi-cellular organisms though split genes are the rule rather than the exception. The whole area between the start and end of the gene is transcribed by the messenger RNA, after transcription an additional step cuts off the DNA which is not needed to make the protein. The mRNA is then spliced back together for making the protein. This work is accomplished by a set of proteins called snRNAs. This process allows the body to re-use the same DNA to make different proteins through alternative splicing. In this manner the human body can make some 100,000 proteins with only some 30,000 genes. There are at least 100 proteins involved in the process of mRNA splicing [1993a]. The discovery of split genes brought Richard Robert and Phil Sharp the [1993]Prize. This came late in the examination of DNA because most early DNA research was done on bacteria which do not have split genes. In multi-cellular organisms though split genes are the rule rather than the exception. The whole area between the start and end of the gene is transcribed by the messenger RNA, after transcription an additional step cuts off the DNA which is not needed to make the protein. The mRNA is then spliced back together for making the protein. This work is accomplished by a set of proteins called snRNAs. This process allows the body to re-use the same DNA to make different proteins through alternative splicing. In this manner the human body can make some 100,000 proteins with only some 30,000 genes. There are at least 100 proteins involved in the process of mRNA splicing [1993a].
It is critical that only fully processed mature mRNAs be exported from the nucleus because translation of incompletely processed pre-mRNAs containing introns would produce defective proteins, which might interfere with the functioning of the cell. By mechanisms that are not fully understood, pre-mRNAs associated with snRNPs in spliceosomes are prevented from being transported to the cytosol. In one type of experiment, for instance, a gene encoding a pre-mRNA with a single intron that is efficiently spliced out was mutated to introduce deviations from the consensus splice-site sequences. Mutation of either the 5â² or 3â² invariant splice site at the ends of the intron resulted in pre-mRNAs that were bound by snRNPs to form spliceosomes; however, RNA splicing was blocked and the pre-mRNA was retained in the nucleus. In contrast, mutation of both the 5â² and 3â² splice sites in the same pre-mRNA resulted in efficient export of the unspliced pre-mRNA. In this case, the pre-mRNAs were not efficiently bound by snRNPs.
[1993b].
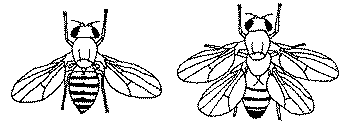 Edward Lewis, Christiane Nusslein-Volhard and Eric F. Wieschaus won the [1995] Prize for discoveries regarding early embryonic development. They looked at the homeobox genes (shortened to Hox genes) of the fly and found that they were arranged in the same sequence as that in which development of the fly occurred. Through the use of induced mutations they established the parts of the fly controlled by each gene. Upon destroying the functioning of one gene, they got a fly with four wings. This was caused by the developmental sequence skipping over the mutated gene and transcribing the following gene in its place. The two extra wings replaced the hateres of the fly which serve as stabilizers in flight, much the same way as the tail wings of a plane serve as stabilizers. The extra wings however were utterly non-functional. Edward Lewis, Christiane Nusslein-Volhard and Eric F. Wieschaus won the [1995] Prize for discoveries regarding early embryonic development. They looked at the homeobox genes (shortened to Hox genes) of the fly and found that they were arranged in the same sequence as that in which development of the fly occurred. Through the use of induced mutations they established the parts of the fly controlled by each gene. Upon destroying the functioning of one gene, they got a fly with four wings. This was caused by the developmental sequence skipping over the mutated gene and transcribing the following gene in its place. The two extra wings replaced the hateres of the fly which serve as stabilizers in flight, much the same way as the tail wings of a plane serve as stabilizers. The extra wings however were utterly non-functional.
This discovery shows several important points. The development of an embryo follows a predetermined programmatic set of steps. That is why when a gene was rendered nonfunctional, the next gene did not do what it normally does. It also shows that a feature, in this case the two new wings, requires more than a single change for it to be useful. The extra wings did not improve on the flying ability of the fly, they decreased it because the hateres served a useful purpose and the new wings were just useless baggage. The following shows that the non-coding DNA is not only an essential part of the development process, but the major part of it:
The DNA sequence of the BX-C has now been completely determined (101) and a preliminary analysis made of it (102). The protein coding regions comprise only 2% of the entire sequence. The other 98% is expected to contain a diverse group of motifs to which trans-regulatory proteins bind, thereby conferring the specific spatial and temporal expression of the protein products of each domain. There may also be a regulatory role for noncoding RNA's of the type identified in the bxd and iab-4 regions.[1995a].
 The [1997] Prize went to Stanley Prusiner for the discovery of the prion, the protein which when folded incorrectly produces mad cow disease and other nervous system disorders. Prion proteins are present in numerous species, but their exact purpose is undetermined as yet. The mutant form of the protein, differing from the normal one only in the way it folds, is very resilient to many agents which normally dissolve other proteins and can withstand extreme heat. For this reason, even cooking meat infected with mutant prions will not kill them, they thus will infect the individual eating it. Prion infection makes the normal prion proteins in the body fold abnormally thus spreading the disease. The [1997] Prize went to Stanley Prusiner for the discovery of the prion, the protein which when folded incorrectly produces mad cow disease and other nervous system disorders. Prion proteins are present in numerous species, but their exact purpose is undetermined as yet. The mutant form of the protein, differing from the normal one only in the way it folds, is very resilient to many agents which normally dissolve other proteins and can withstand extreme heat. For this reason, even cooking meat infected with mutant prions will not kill them, they thus will infect the individual eating it. Prion infection makes the normal prion proteins in the body fold abnormally thus spreading the disease.
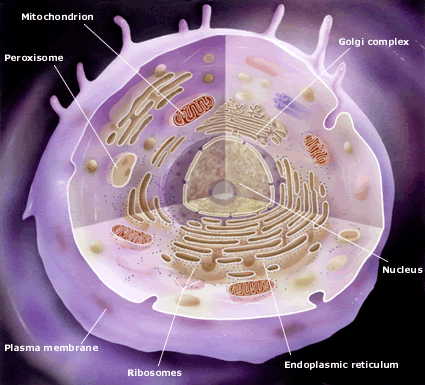
 Gunter Blobel's discovery of the methods behind cell signaling won him the [1999] Prize. The cell itself as well as the nucleus, the mitochondria and other organelles are surrounded by membranes which keep out undesired substances and keep in desired ones. Passage in and out is done through specified channels which only permit passage of desired substances. To achieve passage proteins require a signal sequence of ~15 amino acids, usually at the beginning, which acts as a 'passport' enabling the protein to pass through. " Each signal sequence is membrane specific and is decoded by a complex machinery that is restricted in its location to one particular cellular membrane."[1999a]. Gunter Blobel's discovery of the methods behind cell signaling won him the [1999] Prize. The cell itself as well as the nucleus, the mitochondria and other organelles are surrounded by membranes which keep out undesired substances and keep in desired ones. Passage in and out is done through specified channels which only permit passage of desired substances. To achieve passage proteins require a signal sequence of ~15 amino acids, usually at the beginning, which acts as a 'passport' enabling the protein to pass through. " Each signal sequence is membrane specific and is decoded by a complex machinery that is restricted in its location to one particular cellular membrane."[1999a].
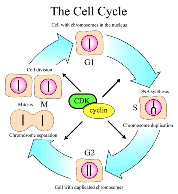 Leland H. Hartwell, R. Timothy (Tim) Hunt and Paul M. Nurse won the [2001]
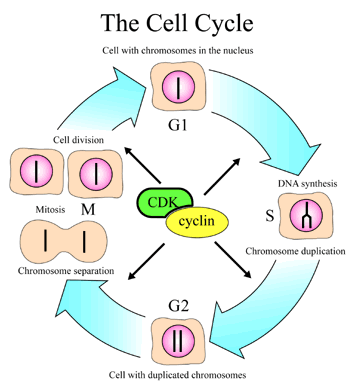
Prize for their discoveries of key regulators of the cell cycle. Hartwell found over 100 genes involved in the control of the cell cycle [2001a]. There are four phases in the cell cycle. In the first the cell starts to grow( G1), in the second it replicates the DNA (S) , it then checks the duplication (G2), the chromosomes separate (M), and lastly the cell divides. At each of these steps there are checkpoints that determine if the previous step has been successfully completed and repairs are made. The steps in the cycle are likewise regulated and occur in the designed order. The two main sets of molecules involved in the cell cycle, CDK and cyclins act as an engine and gears idling or driving the cell cycle forward. This complex system insures proper cell replication which otherwise would lead to cancer and other serious problems. Leland H. Hartwell, R. Timothy (Tim) Hunt and Paul M. Nurse won the [2001]
Prize for their discoveries of key regulators of the cell cycle. Hartwell found over 100 genes involved in the control of the cell cycle [2001a]. There are four phases in the cell cycle. In the first the cell starts to grow( G1), in the second it replicates the DNA (S) , it then checks the duplication (G2), the chromosomes separate (M), and lastly the cell divides. At each of these steps there are checkpoints that determine if the previous step has been successfully completed and repairs are made. The steps in the cycle are likewise regulated and occur in the designed order. The two main sets of molecules involved in the cell cycle, CDK and cyclins act as an engine and gears idling or driving the cell cycle forward. This complex system insures proper cell replication which otherwise would lead to cancer and other serious problems.
At the heart of the G1 checkpoint lies the series of events leading to the accumulation of the tumor suppressor protein, p53. Although p53 exerts a pervasive influence on checkpoint functions during the mammalian cell cycle, the G1 checkpoint represents the only case in which loss of p53 leads to total checkpoint abrogation (Ko and Prives 1996; Giaccia and Kastan 1998; North and Hainaut 2000). DNA damage induced by most, if not all, forms of genotoxic stress induces a rapid increase in the level of p53, a response that is mediated primarily through an increase in protein stability. In addition to triggering the accumulation of p53, genotoxic stress induces posttranslational modifications that regulate the transcriptional activating functions of this protein. With respect to the G1 checkpoint, a key target for transcriptional activation by p53 is the cyclin-dependent kinase inhibitor, p21 (also termed WAF1 or CIP1). The p53-dependent increase in p21 expression suppresses cyclin E and cyclin A-associated cdk2 activities, and thereby prevents G1-to-S phase progression. In addition to p21, the activated form of p53 stimulates the expression of a large panel of genes, which, depending on the cellular context and type of initiating insult, may modulate intracellular redox status, or induce the host cell to undergo apoptosis (Yu et al. 1999).[2001a]
 Sydney Brenner's, H. Robert Horvitz's and John E. Sulston's discovery of genetic development and programmed cell death in the nematode c.elegans won them the [2002] Prize. This small worm has two qualities which made it attractive for developmental research - a small size of only 959 cells and transparency, which made it easy to see its development. One of the surprising discoveries was that during the development cycle the worm destroys exactly 131 cells it had previously made. The studies showed that the developmental process does not vary in any way, all the worms went through the same program of cell division and differentiation and that the death of some cells is specifically programmed. The death genes as well as a gene which prevents cell death were identified. Sydney Brenner's, H. Robert Horvitz's and John E. Sulston's discovery of genetic development and programmed cell death in the nematode c.elegans won them the [2002] Prize. This small worm has two qualities which made it attractive for developmental research - a small size of only 959 cells and transparency, which made it easy to see its development. One of the surprising discoveries was that during the development cycle the worm destroys exactly 131 cells it had previously made. The studies showed that the developmental process does not vary in any way, all the worms went through the same program of cell division and differentiation and that the death of some cells is specifically programmed. The death genes as well as a gene which prevents cell death were identified.
Conclusions:
In all the above we see not randomness, not fortuitousness, not gradualness of system development, but essential functionality, specificity, and complexity. More than half of the above discoveries were made in simple unicellular organisms. The entire systems are clearly essential for even the simplest of living things since they control the essential functions of life, development and reproduction. They are requirements, not fortuitous occurrences and could in no way have 'developed' but had to exist from the beginnings of life. The more advanced organisms likewise require the more advanced systems such as the ones for correctly transcribing split genes, restriction enzymes, cholesterol metabolism, cell signaling, the well regulated immune system and most importantly programmed cell development. Most importantly, each system is not an isolate, but an essential and integral part of the larger system - the living organism. Each new discovery also shows the hopelessness of the undesigned explanations. With each new discovery we find more complexity, more necessary functions, and more proteins required for functions which had already been discovered.
We are easily inclined to hold an exaggerated opinion of ourselves in this era of advanced technology. Thus, we are justified in having a great admiration for the achievements in electronics, where, for example, the attempts at miniaturization to reduce component size, to lower the weight, and reduce the volume of apparatus have enabled a rapid development of space science. However, we should bear in mind that, millions of years ago, nature perfected systems far surpassing all that the inventive genius of man has been able to conceive hitherto. A single living cell, measuring several thousandths of a millimetre, contains hundreds of thousands of chemical control circuits, exactly harmonized and functioning infallibly. It is hardly possible to improve on miniaturization further; we are dealing here with a level where the components are single molecules.[1965]
|
|